

Специфика роста мышц - рнк, гормоны...
| Разместил: 43facog6D | 10 декабря 2008 | Просмотров: 25045 |
Метки: рост мышц, Протасенко, рнк, гормоны
Автор: Вадим Протасенко
Содержание белка в мышцах человека, на первый взгляд, не так велико – от 16% до 21% общей мышечной массы, но если учесть, что 72-80% мышечной массы приходится на воду, то становится ясно, что «сухая» ткань мышц на 75-80% состоит из белка. Из нескольких видов белка, главным образом актина и миозина, состоят миофибриллы – структуры мышечного волокна, выполняющие основную мышечную функцию – функцию сокращения. Цепочки таких белков, как десмин, дистрофин, спектрин и др. формируют каркас мышечных волокон. Белок является главным компонентом различных мембран, как внутриклеточных, так и мембран, составляющих оболочку мышечного волокна. Из белков состоит межклеточная соединительная ткань, связки и сухожилия мышц. Ферменты – вещества, обеспечивающие протекание в мышечной клетке всевозможных химических реакций, связанных с жизнедеятельностью мышц, – это суть тоже белки. Даже внутриклеточная жидкость (саркоплазма) по своей структуре больше напоминает гель, потому как в ней растворено большое количество белка, главным образом все тех же ферментов.
Несмотря на то, что человек получает белки с пищей, белки других живых организмов, потребляемые человеком, не используются напрямую для строительства тканей его тела. Весь поступающий в организм человека белок сначала расщепляется в пищеварительной системе на составные части белка – аминокислоты. В кишечнике аминокислоты просачиваются в кровь и разносятся по всем клеткам организма. И только затем в каждой клетке из поступивших в нее аминокислот собираются белки, характерные для данной клетки данного организма (отличает одни белки от других порядок следования аминокислот в молекуле белка). Синтезированные клетками белки не включаются в клеточные структуры «на века», в тканях организма постоянно происходит обратный распад белков до аминокислот, их составляющих. Часть аминокислот, являющихся продуктами распада белка, расщепляется далее до более простых соединений, но большая часть этих аминокислот, наряду с новыми аминокислотами, поступающими с пищей или синтезированными самим организмом, тут же включается в новые белковые молекулы, встраивающиеся в ткани взамен распавшихся.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Установлено, что за 10 дней обновляется половина всех белков печени и крови человека, немногим дольше живут и мышечные белки, так, известно, что миофибриллярные белки в мышцах кролика полностью обновляются в течение месяца. Таким образом, существование мышечной ткани есть непрерывный процесс обновления белков ее составляющих. Соответственно, от соотношения скоростей распада и синтеза белка зависит то, набирает ли человек мышечную массу или теряет ее. Более того, увеличение силы или выносливости мышц без существенного изменения их массы или объема также связано с накоплением в мышцах определенных видов белка, выполняющих функции обеспечения мышечного сокращения. Так, например, накопление в мышцах окислительных ферментов и миоглобина – белка, осуществляющего внутриклеточный транспорт кислорода, приводит к увеличению скорости воспроизводства энергии за счет окислительных процессов, что в целом увеличивает выносливость мышц. Следовательно, тренировка любой направленности, тренировка «на массу», силовая тренировка, или тренировка выносливости мышц, если она достигает своей цели, приводит к увеличению содержания в мышцах тех или иных видов белка. Точнее будет сказать: увеличение содержания в мышцах определенных видов белка есть причина изменений функциональных свойств мышц в процессе их тренировки.
Потому для понимания путей воздействия тренировки на размер и силовые характеристики мышц важно знать, какого рода тренировка и каким образом способствует накоплению в мышцах тех или иных видов белка.
Влияние тренировки на накопление в мышцах тех или иных видов белка
Теоретически увеличение содержания белка в мышцах возможно как благодаря активизации синтеза белка, так и благодаря снижению скорости его распада. Однако, достоверно установлено, что интенсивная работа мышц активизирует катаболизм белка в мышечной ткани, при этом повышенный уровень распада белка может наблюдаться вплоть до нескольких дней после тренировки. А это, в свою очередь, означает, что увеличение содержания белка в мышцах под воздействием тренировки никак не может быть следствием снижения интенсивности катаболических процессов, следовательно, тренировка должна активизировать синтез белка в большей степени, чем его распад.
Последнее утверждение на данный момент является истиной, не подвергающейся сомнению, тем не менее, сами механизмы воздействия тренировки на процессы синтеза белка в мышцах до настоящего времени в достаточно полной мере еще не изучены и являются предметом дискуссий.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
В очень грубом приближении процесс синтеза белка можно описать нижеследующей схемой.
В каждой клетке человеческого организма, в том числе и мышечной, имеется ядро, внутри которого заключена молекула ДНК. В молекуле ДНК записана информация о строении всех белков организма. Так как отличает один вид белка от другого лишь последовательность аминокислот в аминокислотной цепочке белка, то именно последовательность аминокислот в молекуле того или иного белка закодирована в ДНК. Участок ДНК, содержащий информацию о строении одного вида белка, принято называть геном. При необходимости синтезировать в клетке определенный белок, с гена данного белка снимается особая копия, называемая матричной РНК, затем РНК выходит из ядра в клетку, и далее на РНК как на шаблоне выстраивается молекула белка. Строительство белка осуществляется путем соединения друг с другом свободных аминокислот, имеющихся в клетке, в том порядке, который «записан» в РНК. Молекула РНК используется при строительстве белка не как расходный материал, а как чертеж, план строительства, потому на основе одной молекулы РНК может быть собрано множество молекул белка, но, понятно, что чем больше РНК в клетке, тем большее количество молекул белка может собираться одновременно. К тому же РНК имеет тенденцию со временем распадаться, и для непрерывности синтеза белка требуется постоянное восполнение молекул РНК в клетке.
В итоге интенсивность синтеза того или иного белка в клетке зависит от интенсивности синтеза соответствующей РНК в ядре клетки, то есть, от частоты считывания РНК с гена данного белка. Ядра любой клетки человеческого организма имеют одинаковый набор генов, то есть, содержат информацию обо всех белках организма (порядка 100000 генов), однако большинство генов в клетках неактивно, и лишь на небольшой части генов происходит синтез РНК. Так, в мышечных клетках активируется считывание РНК с генов миозина и актина, генов иных белков, характерных для мышечной клетки, а вот гены других видов белка, например, белков крови или белков соединительной ткани, в клетках мышц «молчат». Да и активность «мышечных» генов, в мышечных клетках также не постоянна и может изменяться в зависимости от условий жизнедеятельности мышц. Тот факт, что свойства мышц под воздействием тренировки могут изменяться, то есть, может изменяться относительное содержание тех или иных видов белка в мышце, свидетельствует о том, что тренировка воздействует именно на механизмы синтеза РНК, активируя считывание РНК с нужных генов. И действительно, во множестве экспериментов отмечено резкое усиление синтеза различных видов РНК в мышечных клетках в первые часы после тренировки.
По-видимому, основываясь на приведенных выше фактах и соображениях, ученые достаточно давно пришли к заключению, что тренировка способствует выработке в мышцах определенных веществ – так называемых факторов-регуляторов, активирующих синтез РНК в ядрах мышечных клеток, благодаря чему после тренировки в мышцах активизируется синтез белка, а при регулярных тренировках наблюдается накопление белков в мышцах, то есть, гипертрофия мышц. Считается, что стероидные гормоны, проникающие в мышечную клетку и соединяющиеся со стероидными рецепторами, воздействуют на ядерную ДНК, активируют синтез РНК некоторых мышечных белков, тем самым усиливая синтез белка в мышцах.
В дальнейшем, я не буду постоянно описывать всю истинную последовательность событий, приводящих к синтезу белковых молекул в клетке. Весь данный процесс, начиная с синтеза РНК в ядре клетки и заканчивая сборкой белковой молекулы в саркоплазме клетки, я для краткости буду называть «синтезом белка ядром клетки», при этом следует помнить, что само ядро вовсе не синтезирует белок, а лишь управляет его синтезом. Данный прием позволит мне более кратко и понятно для читателя сформулировать некоторые важные мысли. Так, в частности, описанную выше схему гипертрофии мышц под воздействием тренировки можно будет заменить следующим кратким утверждением: во время интенсивных мышечных сокращений в мышцах вырабатывается ряд факторов регуляторов, воздействующих на ядра мышечных клеток, что приводит к ускорению «синтеза белка данными ядрами» и в дальнейшем к гипертрофии мышц.
Многие ученые, занимающиеся проблемами спорта (во всяком случае, в России), до сих пор убеждены именно в таком механизме мышечной гипертрофии, что подтверждается как статьями данных ученых, так и содержанием современных учебников по биохимии спорта. Вместе с тем, совокупность накопленных к настоящему времени экспериментальных фактов свидетельствует о том, что представления, согласно которым гипертрофия мышц является следствием интенсификации «синтеза белка ядрами» мышечных клеток, крайне далеки от истинного положения дел.
За счет чего же, на самом деле, растут мышцы?
Так как настоящая статья носит популярный характер, я не буду утруждать себя перечислением, а читателей чтением ссылок на многочисленные исследования, подтверждающие излагаемые мной сведения. Те из читателей, кто останется неудовлетворенным поверхностностью изложения материала в настоящей статье, могут отыскать в сети Интернет на сайте «Проблемы тяжелой атлетики» (http://www.shtanga.kcn.ru) или сайте Федерации пауэрлифтинга России (http://www.russia-pf.ru) мою статью «Функциональная гипертрофия скелетных мышц. Локальные механизмы адаптации скелетных мышц к нагрузке», в которой затронутые в настоящей статье вопросы рассмотрены более подробно с достаточной степенью строгости, а сделанные утверждения подкреплены необходимым анализом и ссылками на соответствующие исследования, проведенные как советско-российскими, так и зарубежными исследователями за последние 40 лет.
Итак, для того, чтобы понять, что что-то не так в традиционных представлениях о механизмах мышечного роста, достаточно задуматься над особенностями строения мышечных клеток и особенностью их формирования в период развития эмбриона. Первым «бросающимся в глаза» отличием мышечных клеток от клеток иных тканей является их размер. Если для того, чтобы увидеть большинство клеток человеческого организма, потребуется микроскоп, то мышечную клетку можно заметить невооруженным глазом. Мышечная клетка – это мышечное волокно, трубчатое образование диаметром, равным примерно толщине человеческого волоса, и длиной от нескольких миллиметров до 12 см (в зависимости от вида мышц и их строения). Образуется эта суперклетка на этапе эмбрионального развития путем слияния большого числа обычных по размеру небольших клеток предшественников (миобластов) в длинные трубчатые структуры (рис. 1). Таким образом, в мышечных клетках-волокнах оказывается не одно ядро, как в других клетках, а множество ядер (как правило, несколько тысяч), по числу клеток эмбриона, слившихся в волокно. Пока в волокне собираются миофибриллы, ядра занимают центральное положение вдоль всей длины волокна, а затем, после окончания формирования волокна ядра, перемещаются к поверхности волокна, где и пребывают в дальнейшем, и откуда управляют синтезом белка. Возникает вопрос, почему мышечное волокно не вырастает из одной клетки, а для его образования требуется слияние столь большого числа клеток? Ответ напрашивается сам собой. По-видимому, одной клетки, точнее, одного ядра совершенно недостаточно для синтеза такого количества белка, которое требуется для формирования и дальнейшего обслуживания столь большой структуры, как мышечное волокно. К тому же, будь в мышечном волокне только одно ядро, даже если бы оно и могло обеспечить синтез белка в неограниченном количестве, то синтезированные белки пришлось бы доставлять от ядра на периферию волокна на слишком большие по молекулярным меркам расстояния. Благодаря же слиянию большого числа клеток воедино, ядра равномерно распределяются вдоль всего мышечного волокна, и объем волокна, который обслуживается одним ядром, оказывается кардинально не отличающимся от объема обычной одноядерной клетки.
Итак, сама многоядерность мышечных волокон свидетельствует о том, что объем мышечного волокна, который способно обслуживать одно клеточное ядро, ограничен.
Известно, что мышцы ребенка, чтобы достигнуть размера, характерного для взрослого человека, должны увеличиться примерно в 20 раз. Если исходить из того, что рост мышц связан с ускорением «синтеза белка ядрами», то следует признать, что по мере взросления человека объем мышечного волокна, обслуживаемый одним ядром, должен увеличиться в его мышцах примерно в двадцать раз, и в такой же пропорции должна возрасти скорость «синтеза белка одним ядром». На самом деле, конечно, ничего подобного не происходит. Исследования, проведенные еще в 70-е годы прошлого века, показали, что объем мышечного волокна, приходящийся на одно ядро, примерно одинаков в мышцах людей в возрасте от 1 года до 70 лет. А это означает, что в мышечных волокнах взрослого человека ядер примерно в 20 раз больше, чем в мышцах ребенка.
Откуда же в мышечных волокнах человека появляются новые ядра?
Оказывается, при образовании мышечных волокон не все клетки эмбриона, из которых развивается мышечная ткань, полностью сливаются с мышечным волокном, часть эмбриональных клеток, примерно 3-10%, оказывается как бы «законсервированными» под оболочкой мышечного волокна (рис. 3). Эти клетки-спутники мышечного волокна получили название клеток-сателлитов или миосателлитоцитов. При получении определенных химических сигналов клетки-спутники высвобождаются из оболочки волокна, интенсивно делятся, затем часть размножившихся клеток снова становится клетками-спутниками, а часть сливается с мышечным волокном, теряя свою оболочку, и ядра клеток-спутников становятся ядрами мышечного волокна. Тем самым в мышечном волокне увеличивается число ядер, способных «синтезировать белок», а вслед за этим увеличивается количество белка в волокне и, соответственно, увеличивается размер мышечного волокна.
Именно деление клеток-спутников и увеличение числа ядер в мышечном волокне, а вовсе не ускорение «синтеза белка существующими ядрами», является причиной гипертрофии мышц по мере роста молодого организма.
Но, может быть, рост мышц за счет деления клеток-спутников происходит только при возрастном росте мышц в длину, а увеличение мышц в диаметре, происходящие вследствие тренировки, не связно с увеличением числа клеточных ядер, и является следствием ускорения «синтеза белка существующими ядрами»? Исследования мышц элитных пауэрлифтеров с экстремально развитой мускулатурой показало, что объем мышечного волокна, приходящийся на одно ядро (то есть, объем волокна, обслуживаемый одним ядром), у спортсменов ничуть не больше, чем у нетренированных людей. А это, в свою очередь, указывает на то, что гипертрофия мышц, вызванная тренировкой, тесно связана именно с увеличением числа ядер в волокне.
Подтверждает этот вывод множество экспериментов, проведенных за последние 25 лет на людях и животных, в которых было напрямую зафиксировано как активирование клеток-спутников, так и увеличение числа ядер в мышечных волокнах после интенсивной нагрузки. В той или иной мере клетки-спутники активируются как после силовых тренировок со штангой, так и после тренировок на выносливость, например, после беговых тренировок или работы на велотренажере. При этом было замечено, что активация клеток-спутников является одной из первых реакций мышечной ткани на нагрузку. Активизация клеток-спутников фиксируется уже через 12-24 часа после перегрузки мышц, а вот существенная гипертрофия мышц наблюдается гораздо позднее, по прошествии дней и даже недель. В защиту устаревших представлений можно было бы предположить, что мышечное волокно под воздействием тренировки сначала увеличивает свой размер за счет интенсификации «синтеза белка существующими ядрами», и только затем, вслед за увеличением объема мышечного волокна, клетки-спутники делятся и добавляют новые ядра в волокно, чтобы восстановить обычную плотность ядер. Факт активации клеток спутников до, а не после гипертрофии мышц опровергает это предположение. Таким образом, с уверенностью можно утверждать, что деление клеток-спутников является причиной гипертрофии мышц, а не ее следствием.
Потенциал роста мышц за счет деления клеток-спутников очень высок. Так, в одном из экспериментов за три месяца перегрузки мышц кошек, число ядер в медленных волокнах мышц увеличилось в два раза, а в быстрых волокнах в 4 раза! Следует отметить, что деление клеток-спутников является не просто важным механизмом мышечной гипертрофии, но обязательным и, по сути, единственным.
Известно, что удаление у животных некоторых мышц приводит к резкому увеличению нагрузки на оставшиеся мышцы, выполняющие сходные функции (мышцы синергисты), что приводит к значительной гипертрофии данных мышц. Оказывается, что если перед удалением части мышц мышцы животных облучить радиацией (радиация нарушает процессы деления клеток-спутников, но не нарушает механизмы синтеза белка), то компенсаторной гипертрофии оставшихся мышц не наблюдается! Это означает, что даже в условиях крайней потребности в увеличении размера мышц, и наличии соответствующих побуждающих стимулов, рост мышц без деления клеток спутников и добавления новых ядер оказывается невозможным!
Влияние тестостерона на синтез белка
Тут у читателя может возникнуть вопрос, а как же обстоит дело в случае применения анаболических стероидов? Ведь известно, что тестостерон ускоряет в мышцах синтез белка, связываясь с соответствующими рецепторами и воздействуя именно на ядра мышечных клеток, ускоряя тем самым «синтез белка ядрами». К тому же известно, что после окончания «стероидной терапии» и сила мышц, и их объем могут существенно снизиться. Возможно, в случае применения анаболических стероидов объем мышечного волокна может изменяться (возрастать при применении стероидов и падать после их отмены) и без изменения числа ядер?
Удивительно, но исследования показывают, что у спортсменов, применяющих содержащие тестостерон препараты, объем мышечного волокна, приходящийся на одно ядро, оказывается ничуть не большим, чем у спортсменов, воздерживающихся от применения данных препаратов, при том, что размер мышц спортсменов, применяющих допинг, значительно превосходит размер мышц «натуральных» атлетов. Из данного факта неизбежно следует вывод – содержащие тестостерон препараты должны способствовать увеличению числа ядер в мышечных волокнах. И действительно, в ряде экспериментов установлено, что мишенью воздействия тестостерона являются именно клетки-спутники, деление которых активизируется под воздействием данного гормона. Так, при инъекционном введении тестостерона людям было зафиксировано увеличение числа ядер в мышечных волокнах, пропорциональное дозе гормона, при этом увеличения объема мышечного волокна, обслуживаемого одним ядром, не только не наблюдалось, но наоборот, объем волокна, приходящийся на одно ядро, уменьшился! Последний феномен можно объяснить только тем, что в момент проведения измерений объем мышечного волокна в мышцах испытуемых еще не успел в достаточной мере увеличиться вслед за стремительно размножившимися ядрами.
Ряд исследователей пришли к заключению, что увеличение секреции тестостерона в период полового созревания юношей и, как следствие, более активное деление клеток-спутников в мышцах юношей является главной причиной столь существенного различия в развитии мускулатуры мужчин и женщин.
Интересно то, что активность деления клеток-спутников зависит от дозировки тестостерона, чем выше доза введенного препарата, тем активнее делятся клетки-спутники. Последнее наблюдение помогает разрешить одно давнее противоречие, известное спортсменам и специалистам в области «химии», о котором мне в очередной раз довелось прочитать совсем недавно в найденной в Сети статье Хосе Антонио. Позволю себе привести цитату из этой статьи:
«Присоединение тестостерона к рецептору клетки рождает командный импульс, который и «запускает» мириады биохимических реакций. Опять-таки, в теории – андрогенных рецепторов у клеток не так и много. Во всяком случае, того тестостерона, который выделяют половые железы, хватает, чтобы занять их все. Тогда откуда же анаболический эффект дополнительного приема андрогенных стероидов? Вот это как раз и непонятно. Тем более, что в медицинском мире бытует упорное мнение о том, что прием искусственных стероидов сокращает в клетках количество андрогенных рецепторов. И наоборот, если уровень тестостерона в крови низок, рецепторов становится больше. Мол, это приспособительная реакция – так клетки увеличивают шанс захвата редких тестостероновых молекул. Между тем, нашлись еретики, которые взялись оспорить данную точку зрения. И что же вы думаете? Есть доказательства, что на практике все обстоит прямо противоположным образом: у кастрированных животных число рецепторов в мышечных клетках падает, а при обратной закачке тестостерона инъекциями – увеличивается, да еще пропорционально количеству гормонов: больше тестостерона – больше рецепторов!»
Загадка, описанная Хосе Антонио, очень легко разрешается, если принять во внимание тот факт, что тестостерон воздействует не только, собственно, на мышечные волокна, но и на клетки-спутники. На мой взгляд, очень даже вероятно, что нормального уровня тестостерона, характерного для взрослого мужчины (или легкого его превышения, вызванного небольшими дозами введенного препарата), действительно, может вполне хватать для заполнения всех (или почти всех) рецепторов тестостерона, имеющихся в мышечных волокнах, и повышение дозы тестостерона сверх определенного уровня не может ускорить «синтез белка ядрами». Во всяком случае, это хорошо согласуется с тем, что сколько-нибудь значительного увеличения объема мышечного волокна, обслуживаемого одним ядром, на практике не наблюдается даже при инъекционном введении тестостерона. Но так как тестостерон может воздействовать не только на рецепторы мышечных волокон, но и на рецепторы клеток-спутников, то это приводит к активированию деления клеток-спутников, и к увеличению числа ядер в мышечном волокне под воздействием гормона. Новые ядра, в свою очередь, генерируют новые рецепторы тестостерона, ведь рецепторы – это тоже белки, и каждое ядро само обеспечивает себя нужным числом рецепторов. Именно синтезом рецепторов вновь образованными ядрами и может быть объяснено «загадочное» появление в мышечных волокнах новых рецепторов после инъекций тестостерона. И именно воздействием тестостерона на активность деления клеток-спутников можно объяснить зависимость анаболического эффекта тестостерона от дозировки гормона, наблюдаемую спортсменами, применяющими стероиды, и упорно отрицаемую рядом теоретиков.
Воздействие гормона роста на гипертрофию мышц
Рассмотрим теперь пути воздействия на гипертрофию мышц другого важнейшего анаболического гормона – соматотропина, или, по-другому, гормона роста. Известно, что инъекционное введение гормона роста или его посредника – IGF-1 (инсулиноподобный фактор роста)[1] способствует гипертрофии мышц, но оказалось, что при применении данных препаратов, так же, как и при применении анаболических стероидов, увеличения объема мышечного волокна, приходящегося на одно ядро, не наблюдается. То есть, главное анаболическое действие гормона роста и IGF-1 заключается в активации деления клеток-спутников.
О том, что гормон роста активизирует размножение почти всех клеток, способных к делению, было известно достаточно давно. Так, например, в любом учебнике эндокринологии можно найти упоминание об активировании деления хрящевых клеток в так называемых зонах роста костей под воздействием гормона роста, за счет чего кости ребенка вырастают в длину. Хорошо известно также о влиянии данного гормона на рост внутренних органов, например, печени и кишечника. Разросшиеся хрящевые ткани на лицах элитных культуристов, особенно женщин, резко бросаются в глаза, поговаривают и об увеличении внутренних органов у бодибилдеров, прибегающих к помощи гормона роста, существует даже такой термин – «гормональный живот». Вместе с тем, воздействие гормона роста на мышечную ткань и ученые, и рядовые спортсмены упорно не желали связывать с клеточным делением, долгое время рассуждая лишь об общем ускорении синтеза белка под воздействием данного гормона. Осознание того факта, что сколько-нибудь существенный рост мышечной ткани возможен только за счет деления клеток-спутников, помогает более детально понять пути, которыми гормон роста приводит к гипертрофии мышц.
Возникает вопрос, а может ли рост мышц происходить не только за счет увеличения объема мышечных волокон, но и за счет увеличения их числа? Не могут ли активированные клетки-спутники сливаться в новые волокна, как это происходит с миобластами при формировании мышц в эмбриональный период? То есть, возможна ли гиперплазия мышечных волокон?
Ничего принципиально невозможного в таком развитии событий нет. В научной литературе неоднократно описаны случаи, когда поврежденные мышечные волокна гибли от полученных повреждений, но клетки-спутники, высвободившиеся из-под оболочки поврежденных волокон, активно делились и затем, сливаясь друг с другом, образовывали новые мышечные волокна взамен утраченных. Наблюдаются такие регенерационные процессы, в том числе, и в мышцах людей, а в экспериментах над животными отмечены не только факты регенерации отдельных волокон, но имеются примеры регенерации целых мышц. Так, если у крыс в условиях стерильности удалить мышцу, измельчить ее и затем измельченную массу уложить обратно в мышечное ложе, то через некоторое время эта биомасса преобразуется в новую мышцу, волокна которой формируются размножившимися клетками-спутниками, высвободившимися из-под оболочки волокон при их измельчении. Конечно, такая мышца после регенерации значительно уступает в размере мышце до повреждений, число волокон в восстановленной мышце оказывается меньшим, чем до операции, а значительная часть мышечной ткани замещается соединительной тканью. Между тем сами такие эксперименты показывают, что клетки-спутники принципиально способны повторить эмбриональный путь развития и образовывать новые мышечные волокна. Такая потенциальная способность клеток-спутников образовывать мышечные волокна оказывается востребованной и в практике физической тренировки мышц. После интенсивной физической нагрузки тонкие новообразованные мышечные волокна обнаруживаются как в мышцах животных, так и в мышцах человека, например, молодые новообразованные волокна были обнаружены в трапециевидных мышцах высокотренированных пауэрлифтеров.
В то же время, обнаружение в мышцах людей и животных молодых развивающихся волокон еще не является свидетельством того, что рост мышц возможен за счет увеличения числа волокон. Достоверно установлено, что интенсивная физическая нагрузка способна приводить к микроповреждениям мышечных волокон (не путать с травмами связанными с разрывом или надрывом связок или мышц), вплоть до полного разрушения части мышечных волокон. В случае таких значительных повреждений волокон клетки чистильщики, живущие в крови и соединительной ткани (нейтрофилы, макрофаги и др.), очищают внутренности волокна от поврежденной ткани, а затем клетки-спутники выстраивают новое мышечное волокно в пределах оболочки старого. Вполне вероятно, что молодые тонкие мышечные волокна, обнаруживаемые в мышцах спортсменов, образуются только взамен сильно поврежденных тренировкой и полностью деградировавших волокон, и увеличения общего числа волокон в результате таких регенерационных процессов не происходит. Вместе с тем, ряд исследователей, зафиксировавших образование молодых волокон в мышцах после перегрузки мышц, склоняется к мнению, что они имеют дело не просто с регенерировавшими волокнами, но именно с образованием новых волокон в межклеточном пространстве, то есть, волокон, дополнительных к уже существующим. Вместе с тем, с уверенностью говорить о гиперплазии мышечных волокон можно только в тех случаях, когда в экспериментах фиксируется именно увеличение числа волокон, а не просто наличие в мышце молодых регенерирующих волокон. И такие эксперименты, во всяком случае применительно к животным, имеются.
Так, было обнаружено увеличение числа волокон в мышцах крыс в первые несколько недель после рождения. Предполагается, что новые волокна в этом случае образуются либо за счет слияния вышедших из мышечного волокна вовне клеток-спутников, либо за счет схожих по морфологии клеток, изначально расположенных не внутри мышечных волокон, а в межклеточном пространстве. Между тем, образование новых волокон в формирующемся организме животных также не может быть достаточным свидетельством того, что рост мышц в зрелом возрасте возможен за счет гиперплазии волокон. Многие из исследователей не смогли обнаружить увеличения числа волокон при гипертрофии мышц, вызванной теми или иными видами перегрузки. В то же время имеется и достаточное количество исследований с обратным результатом. Так, Олвей с соавторами прикреплял груз к одному крылу нелетающих птиц, после месяца ношения груза на крыле число мышечных волокон в нагруженных мышцах птиц оказалось на 51,8 % большим, чем в мышцах с другой – ненагруженной – стороны. Антонио и Гонея, применяя в аналогичных экспериментах прогрессивную нагрузку (со временем увеличивали вес, прикрепленный к крылу), достигли увеличения числа волокон относительно ненагруженной мышцы на целых 82%. Но обращаю ваше внимание на то, что в этих экспериментах ученые имели дело с мышцами птиц, в мышцах млекопитающих достигнутые показатели гиперплазии оказались не столь впечатляющими, однако все равно весьма существенными.
Так, Гонея одним из первых еще в 1977 году обнародовал результаты исследований, в которых была зафиксирована гиперплазия мышечных волокон у млекопитающих. Свои эксперименты ученый проводил на кошках, заставляя их за пищевое вознаграждение поднимать груз одной лапой. После сорока шести недель тренировок мышцы тренированных и нетренированных лап животных были подвергнуты гистохимическому анализу. Общее количество мышечных волокон в тренированных лапах животных было на 19,3% больше, чем в нетренированных. Японский исследователь Тамаки зафиксировал увеличение на 14% числа мышечных волокон в мышцах задних конечностей крыс, регулярно (4-5 раз в неделю) в течение 12 недель выполнявших с помощью специально сконструированного устройства упражнение, аналогичное приседаниям с весом.
Несмотря на успехи в экспериментах с животными, прямых свидетельств увеличения числа мышечных волокон под воздействием тренировок в мышцах человека до сих пор обнаружено не было.
То, что прямые свидетельства гиперплазии волокон в мышцах человека не обнаружены, возможно, связано с ограниченностью применимых к человеку методов функциональной перегрузки мышц и методов оценки числа волокон в мышцах: ведь такие методы функциональной перегрузки, как длительное многодневное растягивание мышц путем подвешивания груза (в наибольшей степени вызывающее гиперплазию волокон у птиц), к человеку применить довольно затруднительно. Существенная гипертрофия мышц человека (как в случае экстремального развития мышц профессиональных бодибилдеров, тяжелоатлетов и пауэрлифтеров) происходит в течение многих лет тренировок; сравнение же числа волокон в мышцах спортсменов до начала тренировок и после многолетнего периода тренировок никогда не проводилось (во всяком случае, мне не известно о таких экспериментах).
Если же проявления гиперплазии волокон у человека имеют ограниченный характер, и она (гиперплазия) вносит существенный вклад в увеличение размера мышц только в накопительном режиме в рамках многолетнего тренировочного периода, то обнаружение проявлений гиперплазии после относительно короткого периода тренировок, ограниченного временными рамками эксперимента, окажется весьма проблематичным – в особенности с учетом ограниченности методов подсчета волокон, применимых к человеку. Эксперименты, в которых у животных было зафиксировано увеличение числа волокон, как правило, сопровождались умерщвлением подопытных животных и подсчетом полного числа волокон в их мышцах. Так, в уже упомянутых экспериментах с птицами и кошками гиперплазия волокон была обнаружена благодаря сравнению полного числа волокон в мышцах, извлеченных из тренированной и нетренированной конечностей одного и того же животного, понятно, что такие прямые методы обнаружения гиперплазии к человеку неприменимы.
Тем не менее, существует по крайней мере одно исследование, опубликованное Сеостром в 1991 г., в котором проявления гиперплазии в мышцах человека исследовались близким методом. Проводил данное исследование, конечно, не врач-садист, а патологоанатом – ученый, занимающийся изучением трупов людей, умерших собственной смертью. Понятно, что при изучении трупов речь не может идти о каком-то сравнении числа волокон «до» и «после» тренировок – идея заключалось в том, чтобы сравнить число волокон в мышцах более развитой (доминирующей) и менее развитой (не доминирующей) конечности одного и того же индивида. Для такого сравнения была выбрана небольшая мышца ноги – anterior tibialis. Оказалось, что мышцы доминирующей опорной конечности (левой для правшей) обладали несколько большим размером и большим числом волокон, при том, что среднее поперечное сечение волокон в мышцах обеих конечностей было одинаковым. Эти наблюдения Сеострома наиболее убедительно свидетельствуют в пользу того, что функциональная гипертрофия мышц человека, возможно, все-таки связана с гиперплазией волокон, хотя тут нельзя исключать и изначальные генетические различия в мышцах доминирующей и не доминирующей конечностей.
В большинстве же случаев об изменении числа волокон у человека под воздействием тренировки приходится судить только на основе косвенных оценок, сделанных путем сопоставления размера мышцы и среднего поперечного сечения волокон в пробах ткани, взятых из данной мышцы. Но результаты даже таких исследований весьма противоречивы.
Например, при сравнении мышц элитных бодибилдеров мужского и женского пола была установлена связь между размером мышцы и числом волокон в ней. Мышцы мужчин имеют в среднем в два раза больший размер, чем мышцы женщин, при этом больший размер мышц мужчин частично объясняется большим поперечным сечением мышечных волокон в их мышцах, но, в то же время, мышцы мужчин имеют и большее число волокон, чем мышцы женщин. Последнее может являться как следствием гиперплазии волокон, так и следствием генетических различий между полами.
В одном из известных мне исследований проводилось сравнение проб, взятых из трицепса элитных культуристов и пауэрлифтеров мирового уровня, с пробами, взятыми из мышц спортсменов, практиковавших тренировки с отягощением только в течение шести месяцев. Оказалось, что, несмотря на большие различия в силе и обхвате рук, между элитными спортсменами и любителями не имелось никакого существенного различия в поперечном сечении мышечных волокон. Эти наблюдения поддерживаются и рядом других исследований, например, было обнаружено, что поперечное сечение волокон в пробах мышечной ткани, взятых из мышц бедра и бицепса бодибилдеров, не отличается от поперечного сечения волокон обычных физически активных людей. Результаты таких исследований указывают на то, что больший объем мышц спортсменов с экстремально развитой мускулатурой связан с большим числом волокон в их мышцах. Объяснение этому феномену можно найти либо в том, что исследуемые спортсмены от природы обладали большим числом волокон, либо в гиперплазии волокон в результате тренировок.
Генетическое объяснение кажется в данном случае наименее убедительным, поскольку из него должно следовать, что изначально исследуемые элитные спортсмены имели очень тонкие волокна и многолетние тренировки смогли привести лишь к тому, что их волокна достигли размера, характерного для обычного среднетренированного человека. Упомянутые наблюдения можно было бы считать надежным свидетельством гиперплазии мышечных волокон у человека, если бы не существование аналогичных исследований, но с противоположным результатом. Так в одном из исследований было выявлено, что элитные культуристы с наиболее развитой мускулатурой, действительно, обладают большим числом мышечных волокон, чем их менее продвинутые коллеги. Но, оказалось, что такой разброс числа волокон наблюдается и у нетренированных людей и ряд нетренированных людей имели такое же число волокон, как и элитные культуристы. Более того, было установлено, что усредненное число волокон в мышцах спортсменов ничуть не больше чем в мышцах не тренированных индивидов, то есть, связи между тренировкой и числом волокон нет. На основании этих фактов исследователи пришли к заключению, что такой параметр как число волокон в мышце определяется, скорее всего, генетически.
Подводя итог сказанному, следует признать, что гиперплазия мышечных волокон у животных возможна, и является следствием повреждения волокон в результате функциональной перегрузки мышц и последующих регенерационных процессов, связанных с активацией клеток-спутников, их активным делением и последующим слиянием в новые мышечные волокна. Возможность же гиперплазии волокон в мышцах человека по-прежнему остается под вопросом.
Возможно, регенерационный потенциал мышц человека не столь велик, чтобы восстановление мышечных волокон после их микротравмирования под воздействием тренировки могло вызвать их гиперплазию, но применение таких стимуляторов клеточного деления, как гормон роста и тестостерон, может значительно повысить регенерационные возможности мышц человека. Вопрос о том, может ли такая фармакологическая интенсификация активности клеток-спутников способствовать образованию новых мышечных волокон в мышцах человека, требует дальнейшего изучения. В последнее время в прессе стали проскакивать сообщения о гиперплазии мышц человека под воздействием гормонов, как об установленном факте, так в четвертом номере за этот год журнала Muscular Development опубликована статья Дэна Гвартни с громким названием «Гиперплазия». Литература, на которую ссылается данный автор, делая свои утверждения об обнаружении факта гиперплазии, мне хорошо знакома – в данных исследованиях речь собственно о гиперплазии мышечных волокон вообще не идет. Похоже, автор статьи просто смешал (уж не знаю по неведению ли, или намеренно, ради сенсационного заголовка) деление клеток-спутников, с гиперплазией волокон. В каком то смысле деление клеток-спутников и увеличение их числа можно назвать гиперплазией, гиперплазией клеток-спутников, но дело в том, что, традиционно, говоря о мышечной ткани, под гиперплазией имеют в виду исключительно увеличение числа мышечных волокон, а не ядер в них. Увеличение же числа волокон у человека, повторюсь, достоверно зафиксировано не было. На данном этапе развития наших знаний о внутримышечных процессах, активируемых тренировкой, нам остается ограничиться только общим, но зато абсолютно достоверным утверждением:
Сколько-нибудь существенная гипертрофия скелетных мышц человека под воздействием регулярной тренировки является следствием деления клеток-спутников и увеличения числа клеточных ядер в мышцах.
Происходит ли увеличение содержания ядер в мышцах только за счет увеличения числа ядер в ранее существовавших волокнах, либо число ядер в мышцах увеличивается еще и за счет ядер вновь образованных мышечных волокон – все это до окончательного разрешения вопроса о возможности гиперплазии мышечных волокон у человека можно специально не обговаривать. Тем более, что для практики спорта возможность или невозможность гиперплазии мышечных волокон принципиального значения не имеет, так как и в том, и в другом случае целью спортсмена, желающего добиться прироста мышечной массы, должно стать стимулирование деления клеток-спутников мышечных волокон. От того, возможна ли гиперплазия мышечных волокон или нет, зависит разве что наша оценка потенциальных возможностей развития мышц человека. Если гиперплазия волокон в мышцах человека имеет место быть – развитие мышц спортсмена фактически ничем не ограничено, в противном же случае у каждого человека существует свой индивидуальный предел мышечного развития, в преодолении которого не способна помочь никакая гормональная терапия.
Почему это так? Чем обусловлен предел мышечного развития в случае невозможности гиперплазии волокон? Не все ли равно за счет чего мышцам увеличивать свой размер – за счет увеличения поперечного сечения мышечных волокон или за счет увеличения их числа? Чтобы понять, в чем заключено принципиальное различие между гипертрофией мышечных волокон и их гиперплазией, достаточно взглянуть на схематическое изображение поперечного разреза мышечного волокна (см. рис. 1).
Как я уже упоминал ранее, ядра мышечных волокон не распределены равномерно по всему объему мышечного волокна, а располагаются по периметру волокна, непосредственно под оболочкой (по-видимому, такое расположение ядер объясняется потребностью в быстром реагировании генетического аппарата мышечных волокон, сосредоточенного в ядрах, на химические сигналы различного рода, поступающие в волокна извне, через его оболочку). Мышечные белки – вещества достаточно нестабильные и требуют постоянной замены, потому для нормального функционирования клеточных структур, расположенных в глубине мышечного волокна, требуется постоянно доставлять туда от ядер либо РНК, либо уже синтезированные белки, и это в условиях плотной упаковки по всему объему волокна миофибрильных нитей, служащих своеобразным барьером на пути транспортируемых веществ. Чем больше мышечное волокно в диаметре – тем большие расстояния вынуждены преодолевать транспортируемые вглубь волокна вещества, и тем труднее в таком волокне обеспечивать своевременную замену белковых молекул в сократительных структурах, расположенных в центральных областях волокна.
Отсюда следует вывод, что должен существовать предельный размер волокна, по достижении которого сколько ни увеличивай число ядер в волокне, а объем волокна возрасти более не сможет, так как ядра оказываются расположенными слишком далеко от внутренних областей волокна, чтобы обеспечить в них нормальный обмен веществ. В случае, если гиперплазия волокон невозможна, то по достижении всеми волокнами своего предельного размера бесполезными окажутся любые тренировки и дозы гормонов. Если же деление клеток-спутников может приводить не только к увеличению числа ядер в существующих волокнах, но и к образованию новых волокон, то в этом случае при росте мышцы критического увеличения транспортных расстояний между ядрами и обслуживаемыми ими клеточными структурами не происходит. Ограничения, накладываемые на размер мышц, в этом случае если и существуют, то не со стороны внутренних механизмов роста мышц, а со стороны остальных систем организма, обеспечивающих жизнедеятельность мышечной ткани.
Если посмотреть на развитие мышц современных элитных бодибилдеров и сравнить их мышцы с мышцами обычных посетителей тренажерных залов, трудно поверить, что тут обошлось без гиперплазии мышечных волокон. С другой стороны, опыт тех же бодибилдеров свидетельствует и о том, что предел мышечного развития все-таки существует, мало кто из спортсменов из года в год штурмующих вершины «Олимпии», демонстрирует прирост именно мышечной массы.
Подводя итог сказанному, я сделал следующее утверждение:
Сколько-нибудь существенная гипертрофия скелетных мышц человека является следствием деления клеток-спутников и увеличения числа клеточных ядер в мышцах.
Те из читателей, кто интересовался теорией, а не только практикой «железного спорта», должны были обратить внимание на тот факт, что данное мое утверждение расходится с общепринятыми представлениями о механизмах воздействия тренировки на синтез белка, как распространенными среди спортсменов, так и доминирующими в учебных пособиях. Если заглянуть в современные учебники, то в большинстве из них можно найти описание примерно следующего механизма воздействия тренировки на белковый синтез (см. рис 1): активная мышечная деятельность приводит к энергетическому утомлению мышц, выражающемуся в снижении содержания в мышцах соединений, служащих источниками быстродоступной энергии (таких как АТФ и креатинфосфат), и накоплению в мышцах продуктов их распада (АДФ, креатина, ортофосфорной кислоты); накопление продуктов энергетического метаболизма, в свою очередь, либо непосредственно, либо опосредовано воздействует на ДНК мышечных клеток, что стимулирует синтез РНК и, как следствие, синтез белка в мышцах (смотрите описание механизма синтеза белка в первой части настоящей статьи).
На первый взгляд может показаться, что описанная в учебниках концепция в корне противоречит сделанным мною выводам о причинах гипертрофии мышц человека. Но, как будет показано далее, противоречие это, действительно, только кажущееся. Дело в том, что механизмы синтеза белка изучались сначала на самых простейших одноклеточных организмах, не имеющих даже ядер, ДНК которых размещается непосредственно в теле клетки (бактерии). Позднее ученые приступили к изучению механизмов регуляции белка в более сложных одноклеточных организмах имеющих ядра, а в настоящее время активно изучается регуляция синтеза белка в многоклеточных организмах. Нетрудно заметить, что в подавляющем большинстве случаев регуляция синтеза белка изучалась в клетках либо совсем не имеющих ядер, либо имеющих только одно ядро. Понятно, что интенсивность синтеза белка в таких клетках может регулироваться только посредством управления интенсивностью синтеза РНК на ДНК данной клетки. Мышечное волокно – это редкое исключение, клетка, имеющая не одно, а множество ядер, да еще и обладающая потенциалом к увеличению их числа. Только в такой клетке возможно усиление синтеза белка не только путем усиления синтеза РНК ядрами клеток, но и путем увеличения числа ядер в клетке. Вот эта потенциальная возможность мышечных клеток увеличивать число ядер и оставалась долгое время без внимания спортивных исследователей, а гипертрофия мышц приписывалась исключительно усилению синтеза РНК мышечными ядрами.
Сейчас, когда мы знаем, что размер мышц увеличивается благодаря увеличению числа ядер в мышцах, не стоит совершать новую ошибку и напрочь забывать о возможности регуляции белкового синтеза посредством управления интенсивностью синтеза РНК в ядрах мышечных клеток, ведь такие механизмы регуляции, как я покажу ниже, играют не менее важную роль в развитии функциональных свойств мышц под воздействием тренировки.
Количество ядер в мышце определяет размер мышцы, но никак не может определять ее «содержание». То, какие мышечные белки заполняют этот заданный числом ядер объем, как раз зависит от того, РНК каких белков и в каком количестве синтезируется ядрами данной мышцы. А вот на интенсивность синтеза РНК тех или иных видов белка как раз и воздействуют внутриклеточные факторы, связанные с режимом функционирования мышцы. И для понимания путей воздействия тренировки на интенсивность синтеза того или иного вида белка как раз и стоит обратиться к результатам многолетних исследований механизмов регуляции синтеза белка в простейших организмах.
Во-первых, данные исследования показали, что регуляция всех видов белка осуществляется по единой принципиальной схеме. Практически для каждого структурного белка в клетках вырабатываются определенные регуляторные белки, которые, соединяясь с рядом веществ, попадающих в клетку извне или образующихся внутри клетки в процессе ее жизнедеятельности, взаимодействуют с ДНК клетки, воздействуют на гены и либо запускают, либо наоборот – блокируют синтез соответствующих белков. Во-вторых, исследования показали, что, несмотря на то, что для каждого вида белка существует индивидуальный набор веществ, инициирующих синтез белка (активаторов синтеза) или блокирующих синтез (ингибиторов синтеза), в том какие именно вещества активируют синтез того или иного вида белка есть некое общее правило.
Вообще говоря, все клеточные белки можно разделить на два больших класса: это белки, используемые в качестве строительного материала клетки, главным образом миофибриллярные (белки, образующие цепочки миофибрилл – структур клетки, осуществляющих ее сокращение), и белки саркоплазмы, главным образом ферменты или, по-другому, энзимы – белки, обеспечивающие протекание в мышечной клетке всевозможных химических реакций, обеспечивающих сократительную активность мышечной клетки да и саму ее жизнь. Содержание ферментов в мышцах доходит до 35% от общей массы мышечного белка. Интересно, что описанный выше механизм регуляции синтеза белка через регуляцию синтеза РНК веществами, являющимися активаторами или ингибиторами синтеза, изучался как раз на примере синтеза ферментов. Было установлено, что в большинстве случаев синтез ферментов активируется при поступлении в клетку веществ, которые перерабатываются под действием данных ферментов.
Если рассмотреть теперь механизмы мышечного сокращения и механизмы восстановления энергии, растраченной при сократительной активности мышц, то окажется, что вещества, накапливающиеся в мышечной клетке в процессе ее сокращения и являющиеся продуктами распада высокоэнергетических веществ, служащих мышце в качестве источников энергии, сами являются исходными веществами для реакций, восстанавливающих запасы энергии в мышечной клетке. Так АТФ (тройной фосфат), отдавая запасенную в ней энергию, распадается на АДФ (двойной фосфат) и свободную фосфатную группу, восстановление запасов АТФ происходит путем обратного присоединения фосфатной группы к АДФ, то есть, АТФ является исходным веществом для химических реакций, восстанавливающих запасы АТФ. Или, например, креатинфосфат – при интенсивном сокращении мышц данное вещество, отдавая запасенную в нем энергию, расщепляется до креатина, а креатин является исходным веществом для получения креатинфосфата. Реакции, восстанавливающие запасы высоко-энергитических веществ, протекают в мышечной клетке в период отдыха после нагрузки, и протекают они под действием специфических ферментов. Синтез же ферментов, как было установлено на опытах с бактериями, активируется веществами, являющимися исходными для химических реакций, активируемых данным ферментом. То есть: расход запасов энергии в мышцах в процессе их сокращения и накопление в мышце продуктов распада энергосодержащих веществ активирует синтез ферментов, обеспечивающих восстановление запасов энергии.
Так сокращение мышц почти в любых режимах приводит к расходу АТФ, что должно активировать выработку ферментов, обеспечивающих восстановление АТФ (в том числе, по-видимому, и ферментов окислительного цикла), а вот синтез креатинкиназы – фермента, восстанавливающего запасы креатинфосфата, должна активировать лишь работа мышц в достаточно интенсивном режиме, способствующем расходу креатинфосфата.
Таким образом, тренировка мышц в том или ином режиме должна активировать синтез ферментов, обеспечивающих восстановление энергии при работе в данном режиме, что при регулярных тренировках должно способствовать накоплению в мышцах соответствующих ферментов и, как следствие, приводить к повышению работоспособности (выносливости) мышц при работе в данном режиме.
Отсутствие же в течение длительного времени работы мышц в определенном режиме означает отсутствие распада энергетических веществ, обеспечивающих данный режим сокращения и, как следствие, отсутствие стимулов к синтезу соответствующих ферментов, а естественный распад синтезированных ранее ферментов приводит к постепенному снижению содержания ферментов в мышцах. Именно по этой причине если мышцу, долгое время не работавшую в определенном режиме, заставить сокращаться в данном режиме, то низкий уровень ферментов не позволит восстанавливать энергию с нужной скоростью, что, естественно, приведет к быстрому утомлению мышцы.
Жизнь ферментов относительно коротка — от нескольких часов до нескольких суток, потому без регулярной активации синтеза ферментов их содержание в мышцах довольно быстро истощается. Вот потому-то в видах спорта, для достижения высоких результатов в которых очень важна выносливость мышц, так важны регулярные и относительно частые тренировки. В практике спорта высших достижений, количество тренировочных занятий может доходить до двух, а в исключительных случаях – и до трех в день. Не уверен, что столь частая нагрузка на мышцы полностью оправдана даже в видах спорта, требующих развития выносливости мышц и соответственно поддержания высокого уровня ферментов в мышцах, но, во всяком случае, для таких видов спорта стремление тренеров к повышению частоты тренировок хотя бы понятно. Но насколько оправдана высокая частота тренировок мышц в бодибилдинге?
Время жизни сократительных белков значительно больше времени жизни ферментов – что-то около месяца против нескольких дней в случае ферментов, но, как я уже неоднократно повторял, объем мышц определяется, в первую очередь, не частотой активации белкового синтеза в мышцах, а числом ядер в мышце. Жизненный срок ядер в точности не установлен, но он составляет никак не часы и сутки, а месяцы и даже годы. Конечно, не следует полагать, что содержание числа ядер в мышцах будет неизменным при бездействии в течение месяцев или даже лет, нет, самоуничтожение ненужных ядер – это вполне естественный процесс, наблюдаемый в длительное время бездействующих мышцах. Но, вместе с тем, частота тренировок, активирующих клеточное деление, требующееся для сохранения и даже развития мышечных объемов, значительно ниже частоты тренировок, активирующих синтез мышечных ферментов, требующихся для поддержания и тем более развития работоспособности мышц.
Это мое утверждение подтверждается самой практикой бодибилдинга. Если еще лет двадцать-тридцать назад в соревновательном бодибилдинге тренировка каждой мышцы 3 раза в неделю была практически общепринятой нормой, то сегодня считается допустимым нагружать одну мышечную группу не чаще одного раза в 4-8 дней, и мышечные объемы выступающих атлетов от этого нисколько не уменьшились. Все верно, выносливость, работоспособность мышц и мышечные объемы – это далеко не одно и то же. Но, вместе с тем, насыщенность мышц ферментами, все-таки, в некоторой степени влияет на объем мышц.
Дело в том, что основной объем мышц создает вовсе не собственно белок, а вода, которая удерживается белками внутри мышечных волокон. Белки вместе с липидами создают каркас мышечного волокна, ее оболочку, но наполняет волокно объемом именно вода. Мышечное волокно окружено оболочкой, которая представляет собой растягивающуюся мембрану с избирательной способностью пропускать те или иные виды веществ. Вода может свободно циркулировать сквозь такую мембрану туда и обратно, задавая объем мышечного волокна. А определяет то, какой объем воды находится внутри мышечного волокна, количество растворенных в воде внутри мышечного волокна веществ, всевозможных ионов и, в том числе, ферментов. Чем больше растворенных веществ внутри мышечного волокна, тем больше воды проходит внутрь клеточного волокна сквозь оболочку и тем сильнее «раздувается» мышечное волокно. Все скоротечные эффекты, выражающиеся в увеличении объема мышц уже во время тренировки или в первые несколько дней после нее связаны вовсе не с усилением белкового синтеза, а с накоплением в мышце продуктов распада энергетических веществ (а в определенных случаях и самих внутренних структур клетки). Увеличение в мышечных волокнах растворимых веществ вызывает мгновенный приток воды в волокна, в результате чего мышечные волокна, а вслед за ними и мышцы раздуваются на глазах.
Но вернемся к ферментам. Насыщенность мышечных волокон ферментами не может не сказаться на объеме мышц, чем больше ферментов в мышечных волокнах, тем объемнее будут мышцы при том же количестве ядер и сократительных элементов. Но у насыщенности мышц ферментами есть определенный предел, и сколько ни учащай тренировки, после достижения определенного состояния мышц дальнейшего увеличения количества ферментов в мышцах наблюдаться не будет, а, следовательно, не будет и никакого роста мышц без появления в мышцах новых ядер. Такая картина отсутствия роста мышц при вроде бы регулярных тренировках хорошо знакома спортсменам под названием «тренировочное плато» (не следует путать с состоянием перетренированности). По достижении определенной кондиции мышц прирост результатов и объемов мышц прекращается без стрессовых тренировок, активирующих деление клеток-спутников в мышцах.
Итак, воздействие тренировки на мышцы реализуется не каким-то одним, а, как минимум, двумя альтернативными путями, и схему влияния тренировки на механизмы синтеза белка, взятую мной из современных учебников по биохимии спорта (см. рис.1) стоило бы заменить на более полную (см. рис. 2).
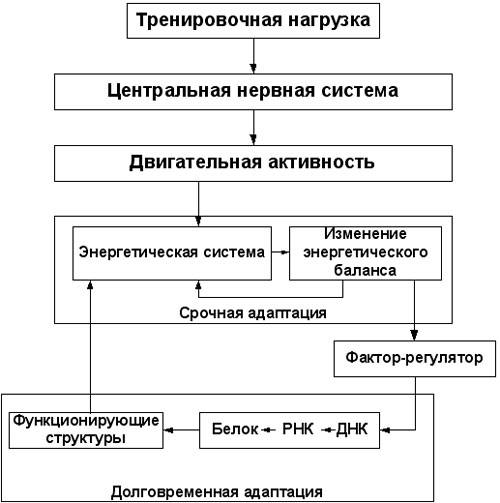
Рис. 1. (схема взята из учебного пособия «Биохимия мышечной деятельности» Н.И. Волков, Э.Н. Несен, А.А. Осипенко, С.Н. Корсун. Издательство «Олимпийская литература» 2000 г.)
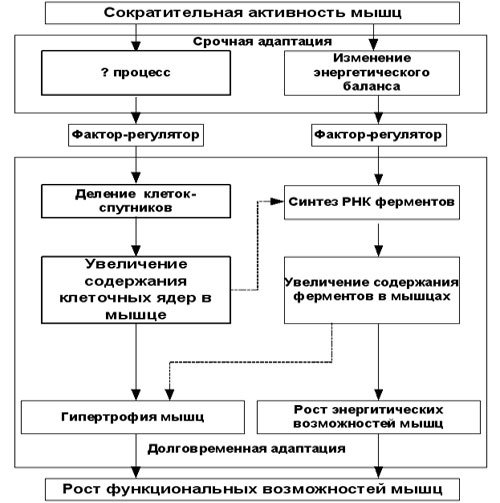
Рис. 2.
В приведенной схеме причина активации клеток-спутников помечена знаком вопроса. Не то, чтобы на современном этапе развития науки факторы эти вообще не были известны ученым, просто современные знания в этой области пока не позволяют сделать однозначные заключения. Достоверно установлено, что активирование клеток-спутников осуществляется факторами, вырабатывающимися при повреждении мышц – так реализуется механизм мышечной репарации. Повреждение мышц тут же вызывает активацию клеток-спутников, что обеспечивает резкое увеличения синтеза белка в мышечной клетке, белка, необходимого для «латания» повреждений. Но вот является ли данный механизм доминирующим при тренировке мышц, и уж тем более – является ли данный путь единственным путем активации клеток-спутников – вот вопрос, на который пока нет точного ответа.
Наверное, всем читателям известно бытующее между спортсменами выражение «без боли нет роста». А ведь боль – это сигнал повреждения мышц, так что можно утверждать, что опыт бодибилдинга, сконцентрированный в этом выражении, показывает, что с высокой вероятностью рост мышц в результате активации клеток-спутников можно связывать с микроразрушениями мышц, полученными в ходе тренировки. Вместе с тем, нужно отдавать себе отчет в том, что это пока только, что называется, «рабочая гипотеза».
Посмотрим же, какие выводы об основных принципах планирования тренировочной нагрузки следуют из достоверно установленных механизмов гипертрофии мышц, сведенных в единую схему на рисунке 2, и данной рабочей гипотезы.
Тяжелые нагрузки, разрушающие мышечную ткань и активирующие деление клеток-спутников, требуют длительного восстановления. Так, только очищение мышц от поврежденной ткани после тяжелых нагрузок может продлиться несколько суток, о чем свидетельствует пик боли, наступающий в мышцах на вторые сутки после тренировки, а пик боли как раз и есть пиком саморазрушения поврежденной при тренировке ткани. Еще около суток требуется на один цикл деления клеток-спутников, это деление следует за самоочищением мышц от повреждений (а таких циклов после одной тренировки может быть несколько). К этому времени стоит еще добавить время, требующееся на синтез РНК новообразованными ядрами и строительство белка, необходимого для воссоздания объема клетки, соответствующего каждому ядру в нормальном неповрежденном мышечном волокне. Следовательно, интенсивные нагрузки, сопровождающиеся мироразрушениями мышечной ткани, не должны применяться слишком часто. Судя по всему, такие нагрузки допустимы не чаще одного раза в неделю на одну мышечную группу, а то и реже. В то же время для поддержания высокого уровня ферментов в мышцах тренировки одной мышцы должны осуществляться гораздо чаще.
Какие способы разрешения данного противоречия существуют? При всем богатстве выбора возможных вариантов, в большинстве своем уже реализованных в тренировочных методиках тех или иных практиков, для бодибилдинга я бы выделил два основных подхода.
Первый подход заключается в разделении годового тренировочного плана на периоды (мезоциклы) различной направленности. В самом простом случае можно выделить лишь два таких мезоцикла – назовем их «базовым» и «предсоревновательным».
В базовый период должны применяться тяжелые тренировки, стимулирующие микроповреждение ткани мышц и последующие регенерационные процессы в мышцах. Такие тренировки требуют длительного отдыха после нагрузки, а потому должны проводиться достаточно редко, не чаще раза в неделю для каждой мышцы (возможно и реже, в зависимости от индивидуальных восстановительных способностей спортсмена, а также в зависимости от применяемых или не применяемых фармакологических препаратов). К окончанию базового периода перед спортсменом стоит задача увеличить количество клеточных ядер в мышцах, и, следовательно, «базовый», связанный с числом ядер объем мышц.
Задача предсоревновательного периода должна заключаться в том, чтобы «выжать» максимум из достигнутого «базового» объема мышц путем еще большего увеличения мышечного объема за счет наполнения мышечных волокон ферментами, и, как следствие, водой (воду внутри мышечных волокон, которая на самом деле содержится там в виде геля из-за высокой концентрации растворенных в ней веществ, не стоит путать с водой в межклеточном пространстве, от которой перед соревнованиями спортсмены как раз стремятся избавиться). Потому в предсоревновательный период интенсивность тренировок должна быть снижена, а частота тренировок увеличена до нескольких тренировок на одну мышцу в неделю. В этот период повреждения мышц при тренировке противопоказаны, ибо это увеличивает требования к времени восстановления мышц, а перед спортсменом стоит задача только довести мышцы до умеренного энергетического истощения, чем стимулировать синтез ферментов в мышцах. Увеличение частоты тренировок и общих энергозатрат организма как нельзя лучше способствует достижению еще одной стоящей перед спортсменом задачи – снижению перед соревнованиями содержания подкожных жиров.
Существует и второй принципиальный подход к тренировкам, позволяющий в одной тренировочной программе совместить требования к длительному восстановлению мышц после тяжелых тренировок, вызывающих разрушение мышечной ткани и деление клеток-спутников, с требованиями высокой частоты тренировочных занятий для насыщения мышц ферментами. Этот подход базируется на введении в тренировочную практику тренировочных занятий различной направленности и интенсивности и затем циклировании таких тренировочных занятий. Основное правило тут следующее: после одного или нескольких тяжелых интенсивных занятий должно следовать несколько более легких тренировок, не вызывающих разрушения мышечной ткани и не мешающих восстановительным процессам. Особенностью таких методик является то, что при их использовании не наблюдается ярко выраженных пиков спортивной формы и ее снижения, как при первом рассматриваемом подходе, что может быть полезным при желании поддерживать достаточно высокую спортивную форму в течение всего годового периода. Но, на мой взгляд, такой подход к тренировкам увеличивает требования к восстановительным возможностям организма.
На этом я, пожалуй, завершу свой экскурс в биохимию и физиологию мышечной деятельности. Надеюсь, что изложенные в данной статье сведения помогут вам, уважаемые читатели «Железного мира», лучше ориентироваться в многообразии существующих тренировочных систем и методик, и дадут вам в руки критерий, позволяющий выбрать среди этого многообразия методику, наиболее соответствую стоящим перед вами целям и имеющимся у вас возможностям.
Успехов вам в достижении целей.
[1] Гормон роста воздействует на рецепторы, расположенные в оболочке клеток печени, мышечной ткани, и клеток ряда других тканей, что приводит к выработке данными клетками инсулиноподобного фактора роста (IGF-1), и уже именно IGF-1 оказывает на ткани организма активизирующее рост воздействие. Выбросы в кровь гормона роста наблюдаются несколько раз в сутки и уровень данного гормона в крови подвержен сильным колебаниям, в то время как уровень IGF-1 значительно более устойчив, и в целом зависит от общего выброса гормона роста.
Сейчас, когда мы знаем, что размер мышц увеличивается благодаря увеличению числа ядер в мышцах, не стоит совершать новую ошибку и напрочь забывать о возможности регуляции белкового синтеза посредством управления интенсивностью синтеза РНК в ядрах мышечных клеток, ведь такие механизмы регуляции, как я покажу ниже, играют не менее важную роль в развитии функциональных свойств мышц под воздействием тренировки.Тренировка мышц в том или ином режиме должна активировать синтез ферментов, обеспечивающих восстановление энергии при работе в данном режиме, что при регулярных тренировках должно способствовать накоплению в мышцах соответствующих ферментов и, как следствие, приводить к повышению работоспособности (выносливости) мышц при работе в данном режиме. Конечно, не следует полагать, что содержание числа ядер в мышцах будет неизменным при бездействии в течение месяцев или даже лет, нет, самоуничтожение ненужных ядер – это вполне естественный процесс, наблюдаемый в длительное время бездействующих мышцах. Но вместе с тем частота тренировок, активирующих клеточное деление, требующаяся для сохранения и даже развития мышечных объемов, значительно ниже частоты тренировок, активирующих синтез мышечных ферментов, требующихся для поддержания и тем более развития работоспособности мышц.Все скоротечные эффекты, выражающиеся в увеличении объема мышц уже во время тренировки или в первые несколько дней после нее связаны вовсе не с усилением белкового синтеза, а с накоплением в мышце продуктов распада энергетических веществ (а в определенных случаях и самих внутренних структур клетки). Увеличение в мышечных волокнах растворимых веществ вызывает мгновенный приток воды в волокна, в результате чего мышечные волокна, а вслед за ними и мышцы раздуваются на глазах. Наверное, всем читателям известно бытующее между спортсменами выражение «без боли нет роста». А ведь боль – это сигнал повреждения мышц, так что можно утверждать, что опыт бодибилдинга, сконцентрированный в этом выражении, показывает, что с высокой вероятностью рост мышц в результате активации клеток-спутников можно связывать с микроразрушениями мышц, полученными в ходе тренировки. Вместе с тем нужно отдавать себе отчет в том, что это пока только, что называется, «рабочая гипотеза». Задача предсоревновательного периода должна заключаться в том, чтобы «выжать» максимум из достигнутого «базового» объема мышц путем еще большего увеличения мышечного объема за счет наполнения мышечных волокон ферментами, и, как следствие, водой (воду внутри мышечных волокон, которая на самом деле содержится там в виде геля из-за высокой концентрации растворенных в ней веществ, не стоит путать с водой в межклеточном пространстве, от которой перед соревнованиями как раз стремятся избавиться).
